



Bulletin E2994
Potato Diseases: Rhizoctonia Stem Canker and Black Scurf
DOWNLOAD
October 23, 2015 - Phillip Wharton
 Print
Print Email
EmailIntroduction
Rhizoctonia disease of potato are caused by the fungus Rhizoctonia solani Kuhn (teleomorph Thanatephorus cucumeris [A. B. Frank] Donk) and can be found on all underground parts of the plant at various times during the growing season. Rhizoctonia solani has many synonyms and is divided into subgroups called anastomosis groups (AGs), in which isolates are categorized according to the ability of their hyphae to anastomose (fuse) with one another. Three AGs of R. solani are prevalent in Michigan: AG2-2, AG-3 and AG-4. AG-3 is the causal agent of disease in potato; AG2-2 and AG-4 cause sugar beet crown and root rot and seedling damping-off, respectively. Rhizoctonia solani AG-3 is relatively specific to potato, and sclerotia on tubers belong almost exclusively to AG-3. Other AGs of R. solani, such as AG2-2 and AG-4, may be pathogenic to potato at some temperatures, but they generally cause little damage.
In Michigan, R. solani causes black scurf on tubers (Fig. 1) and stem and stolon canker on underground stems and stolons (Fig. 2), and it occurs wherever potatoes are grown. However, R. solani causes economically significant damage only in cool, wet soils. In the more southern temperate areas of Michigan, losses from Rhizoctonia are sporadic and occur only when the weather is cold and wet in the weeks following planting. In northern areas, where growers often must plant in cold soils, Rhizoctonia is a more consistent problem. Poor stands, stunted plants, reduced tuber number and size, and misshapen tubers are symptoms of the disease area found on both above- and below-ground portions of the plant. Black scurf (Fig. 1) is the most conspicuous sign of Rhizoctonia disease. In this phase of the disease, the fungus forms dark brown to black, hard masses on the surface of the tuber. These are called sclerotia and are resting bodies of the fungus (Fig. 1). Sclerotia are superficial and irregularly shaped, ranging from small, flat, barely visible blotches to large, raised lumps. Although these structures adhere tightly to the tuber skin, they do not penetrate or damage the tuber, even in storage. However, they will perpetuate the disease, and if infected tubers are used as seed, inhibit the establishment of potato plants.

Figure 1. Rhizoctonia solanisclerotia on the surface of tubers.

Figure 2. Brown, sunken lesions on underground stems and stolons are caused by R. solani.
Although black scurf is the most noticeable sign of Rhizoctonia, stem canker (Fig. 2) is the most damaging component of the disease. It occurs underground and often goes unnoticed. Early in the season, the fungus attacks germinating sprouts underground before they emerge from the soil (Fig. 3). Sprouts may be killed outright if lesions form near the growing tip (Fig. 3). Damage at this stage results in delayed emergence and is expressed as poor and uneven stands with weakened plants. Reduction in crop vigor results from expenditure of seed energy used to produce secondary or tertiary sprouts to compensate for damage to primary sprouts. Occasionally, heavily infested potato seed tubers are unable to produce stems. Instead, the tubers will produce stolons with several small tubers. This symptom, referred to as “no top”, can be confused with the same symptom caused by physiologically old seed that has been de-sprouted.
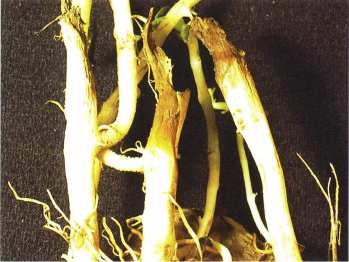
Figure 3. Germinating sprouts may be killed by R. solani before they emerge from the soil.
Poor stands may also be mistaken for seed tuber decay caused by Fusarium for seed tuber decay caused by Fusarium or soft rot bacteria unless plants are dug up and examined. Rhizoctonia does not cause seed decay, damaging only sprouts and stolons. Poor stands and stunted plants can also be caused by blackleg, a bacterial disease that originates from seed tubers and progresses up stems, causing a wet, sometimes slimy rot. In contrast, Rhizoctonia lesions are always dry and usually sunken. Stolons and roots can also be infected by R. solani. Early in disease development, stolons, roots and stems have reddish brown to brown lesions. As red to brown lesions mature, they become cankers that are rough and brown and can have craters, cracks or both. Damage varies and can be limited to a superficial brown area that has no discernible effect on plant growth to severe lesions that are large, sunken and necrotic. If cankers are severe, they may girdle the stem, interfering with the normal movement of water and carbohydrates throughout the plant.
Late-season damage to plants is a direct result of cankers on stolons and stems causing problems with starch translocation. Stolon cankers also affect the shape, size and numbers of tubers produced. If stolons and underground stems are severely infected, the flow of starch from the leaves to the developing tubers is interrupted. This results in small, green tubers, called aerial tubers, forming on the stem above the soil (Fig. 4). Formation of aerial tubers may indicate that the plant has no tubers of marketable quality belowground. Interruptions in carbohydrate flow may also result in a stunting or resetting of the plant. A leaf curl, which can be confused with symptoms of the potato leaf roll virus, has also been reported in severely infected plants.

Figure 4. Small aerial tubers may form aboveground if stolons and underground stems are severely infected.
Disease cycle
Rhizoctonia diseases are initiated by seed-borne or soil-borne inoculum. The pathogen overwinters as sclerotia and mycelium on infected tubers, in plant residue or in infested soils (Fig. 5). When infected seed tubers are planted in the spring, the fungus grows from the seed surface to the developing sprout, and infection of root primordia and leaf primordia can occur. Seed-borne inoculum is particularly effective in causing disease because of its close proximity to developing sprouts and stolons.
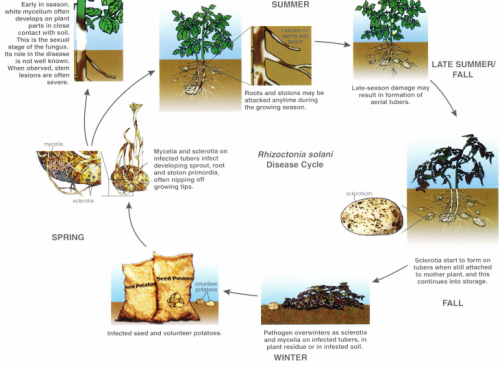
Figure 5. The disease cycle of the stem canker and black scurf pathogen, Rhizoctonia solani.
Mycelia and sclerotia of R. solani are endemic to Michigan soils, living on organic debris, and can cause disease independently of or in conjunction with seed-borne inoculum. Soil-borne inoculum is potentially as damaging as seed-borne inoculum, but it can cause infection only when the plant organs develop in proximity to the inoculum. Roots and stolons may be attacked anytime during the growing season, although most infections probably occur in the early part of the plant growth cycle. The plant’s resistance to stolon infection increases after emergence, eventually limiting expansion of lesions.
Previous research has shown that soil temperature is a critical factor in the initiation of Rhizoctonia disease in potato, with disease severity being positively correlated with the temperature that is most favorable for pathogen growth. The temperature range for the growth of R. solani AG-3 is 41 to 77 degrees F, so plants will be most susceptible to infection when the soil temperatures are within this critical range. Cool temperatures, high soil moisture, fertility and a neutral to acid soil (pH 7 or less) are thought to favor development of Rhizoctonia diseases of potato. Damage is most severe at cool temperatures because of reduced rates of emergence and because growth of stems and stolons is slow relative to the growth of the fungus. Wet soils warm up more slowly than dry soils. This exacerbates damage because excessive soil moisture slows plant development and favors fungal growth. It has been shown that high soil temperatures, especially during the early stages of plant development, tend to minimize the impacts of R. solani, even when inoculum is abundant.
Sclerotia begin to form late in the season, principally after vine death. The mechanisms involved in sclerotial development on daughter tubers are different from those acting in the infection of the mother plant. The mechanisms that trigger sclerotial formation are not well understood, but they may involve products related to plant senescence. However, daughter tubers produced from infected mother plants do not always become infested with sclerotia.
Monitoring and control
Currently, it is not possible to completely control Rhizoctonia diseases, but following a combination of cultural and crop protection strategies may limit their severity. Effective management of this disease requires implementation of an integrated disease management approach and knowledge of each stage of the disease. Although the most important measures are cultural, chemical controls should also be utilized. To date, there have been no comparisons of the relative susceptibility of potato varieties currently grown in Michigan.
Cultural control
One of the keys to minimizing disease is to plant certified seed free sclerotia. If more than 20 sclerotia are visible on one side of washed tubers, consider using a different seed source. Tuber inoculum is more important than soil inoculum as the primary cause of disease. Seed growers should plant only sclerotia-free seed.
Following practices that do not delay emergence in the spring minimizes damage to shoots and stolons and lessens the chance for infection. Planting seed tubers in warm soil (above 46 degrees F) and covering them with as little soil as possible speeds sprout and stem development and emergence, and reduces the risk of stem canker. Plant fields with coarse-textured soils first because they are less likely to become water-logged and will warm up faster.
Rhizoctonia does not compete exceptionally well with other microbes in the soil. Increasing the rate of crop residue decomposition decreases the growth rate of Rhizoctonia. Residue decomposition also releases carbon dioxide, which reduces the competitive ability of the pathogen. The fungus is not an efficient cellulose decomposer, so soil populations are greatly reduced by competing microflora and less disease is observed.
Potatoes should be harvested as soon as skin is set so minimal bruising will occur. The percent of tubers covered with sclerotia increases as the interval between vine kill and harvest increases. Vine removal or burning also reduces the amount of fungus overwintering and thus the amount of inoculum available to infect future potato crops. Do not dump infested tubers on future potato fields as they can become sources of inoculum.
Biological control
There is growing evidence that a biofumigation treatment from incorporating a mustard cover crop is one way to reduce Rhizoctonia incidence. When incorporated into the soil, mustard residues release cyanide-containing compounds that fumigate the soil. At the same time, they also release carbon and nutrients that are the feedstock for soil organisms. Incorporating green cover crop tissues provides energy that supports the complex web of soil organisms that compete with parasite and disease organisms. Thus, mustards and related brassica plant species such as oil-seed radish do not leave a soil void of organisms. Instead, these cover crops tend to tip the balance in favor of beneficial organisms and against parasites and pests.
Our preliminary research indicates that it is important to maximize growth of the cover crop using a high seed rate (15 pounds per acre or more) and irrigate to improve establishment if rainfall is insufficient. A tiny seed such as mustard cannot be drilled too deep. It appears to establish well if broadcast and harrowed or irrigated into sandy soil. The biofumigation benefits of mustard residues are maximized if they are incorporated at or just before flowering. We suggest that residues be mowed and incorporated while still green. Mustards grow rapidly and can become weeds in a subsequent crop, so it is important not to let this cover crop produce seed (see Cover Crop Choices for Michigan Vegetables, MSU Extension bulletin E2896, http://web4.msue.msu.edu/veginfo).
We are just beginning to understand the exact mechanisms involved in biofumigation using mustard cover crops. Initial results from Michigan research indicate that Oriental mustard can be used as a cover crop to improve potato root and tuber health. The growth of Rhizoctonia was slowed by 90 percent in soil amended with Oriental mustard cover crop tissue compared with bare soil. A field experiment indicated that tubers of the table-stock variety Onaway had no observable signs of Rhizoctonia when grown after a spring cover crop of Oriental mustard. Further research is required to learn more about management practices that optimize the biofumigation action of mustard cover crops, but initial results are promising, and farmers are encouraged to experiment with brassica cover crops such as Oriental and white mustard or oil-seed radish to improve soil health.
Chemical control
Seed treatment
Several products have been developed specifically for control of seed-borne potato diseases (Table 1) and offer broad-spectrum control for Rhizoctonia, silver scurf, Fusarium dry rot, and to some extent, black dot (Colletotrichum coccodes). These include Tops MZ, Maxim MZ (and other Maxim formulations + mancozeb) and Moncoat MZ. The general impact of these seed treatments is noted in improved plant stand and crop vigor, but occasionally, application of seed treatments in combination with cold and wet soils can result in delayed emergence. The delay is generally transient and the crop normally compensates. The additional benefit of including mancozeb is prevention of seed-borne late blight.
Table 1. Product name, active ingredient and FRACa resistance management grouping, type and rate of application, and activity of products currently registered for control of Fusarium dry rot of potatoes.
|
Productb |
Active ingredient [Chemical group] |
Type of application |
Rate of applicationc |
Activity against dry rotd |
|
Quadris |
Azoxystrobin (22.9%) [11] |
In-furrow |
6.2-15.4 fl. Oz/A |
+++ |
|
Moncut 70DF |
Flutolanil (70%) [7] |
In-furrow |
0.7-1.1 lb/A |
+++ |
|
MonCoat MZ |
Flutolanil (1.5%) [7] Mancozeb (6%) [M3] |
Seed treatment (dry) |
0.75-1.0 lb/100/b |
+++ |
|
Maxim 4FS |
Fludioxinil (40.3%) [12] |
Seed treatment (liq.) (for production of certified seed tubers only) |
0.08-0.16 fl oz/100lb |
+++ |
|
Maxim Potato Seed Protectant |
Fludioxinil (0.5%) [12] |
Seed treatment (dry) |
0.5 lb/100lb |
+++ |
|
Maxim MZ |
Fludioxinil (0.5%) [12] Mancozeb (6%) [M3] |
Seed treatment (dry) |
0.5 lb/100lb |
+++ |
|
Nubark Maxim |
Fludiozxinil (0.5%) [12] |
Seed treatment (dry) |
0.5 lb/100 lb |
+++ |
|
Tops MZ |
Thiophanate-methyl (2.5%) [1] Mancozeb (6%) [M3] |
Seed treatment (dry) |
0.75 lb/100 lb |
+++ |
a Fungicide Resistance Action Committee, See (http://www.frac.info) for more information.
b Specific instructions are included on the labels of all of the products, and these must be adhered to.
c Rate of application is per acre at 34-inch spacing between rows (in-furrow) and per 100 lb of tuber seed pieces (seed treatment, wet or dry).
d + signifies excellent activity against seed-borne Rhizoctonia.
In-furrow fungicide application
In-furrow application of fungicide at planting has resulted in significant improvement in control of Rhizoctonia disease of potatoes. Products such as Moncut and Quadris applied in-furrow at planting have given consistent and excellent control of Rhizoctonia diseases of potatoes in trials at MSU. However, both seed treatments and in-furrow applications have on some occasion resulted in poor control of Rhizoctonia. This sporadic failure may be due to extensive periods of wet and cold soil shortly after planting or planting in fields with plentiful inoculum. Quadris applied in-furrow has been reported to reduce the symptoms of black dot on lower stems and tubers.





